sábado, 12 de junio de 2010
jueves, 3 de junio de 2010
PROBLEMAS DE GENÉTICA MENDELIANA
Páginas en las que podeis encontrar muchos problemas de genética mendeliana con su soluciones.
En esta página se explican las soluciones paso a paso.
http://www.bioygeo.info/Problemas_gen2.htm
Esta página contiene una gran cantidad de problemas agrupados por categorías.
http://www.biologia.arizona.edu/mendel/mendel.html
Más problemas resueltos.
http://go2.wordpress.com/?id=725X1342&site=matragut.wordpress.com&url=http%3A%2F%2Fmatragut.files.wordpress.com%2F2010%2F03%2Fproblemas-de-genetica-resueltos-2.doc&sref=http%3A%2F%2Fmatragut.wordpress.com%2F2010%2F03%2F18%2Fproblemas-de-genetica-2%2F
http://go2.wordpress.com/?id=725X1342&site=matragut.wordpress.com&url=http%3A%2F%2Fmatragut.files.wordpress.com%2F2010%2F03%2Fproblemas-de-genetica-resueltos-4.pdf&sref=http%3A%2F%2Fmatragut.wordpress.com%2F2010%2F03%2F18%2Fproblemas-de-genetica-2%2F
En esta página se explican las soluciones paso a paso.
http://www.bioygeo.info/Problemas_gen2.htm
Esta página contiene una gran cantidad de problemas agrupados por categorías.
http://www.biologia.arizona.edu/mendel/mendel.html
Más problemas resueltos.
http://go2.wordpress.com/?id=725X1342&site=matragut.wordpress.com&url=http%3A%2F%2Fmatragut.files.wordpress.com%2F2010%2F03%2Fproblemas-de-genetica-resueltos-2.doc&sref=http%3A%2F%2Fmatragut.wordpress.com%2F2010%2F03%2F18%2Fproblemas-de-genetica-2%2F
http://go2.wordpress.com/?id=725X1342&site=matragut.wordpress.com&url=http%3A%2F%2Fmatragut.files.wordpress.com%2F2010%2F03%2Fproblemas-de-genetica-resueltos-4.pdf&sref=http%3A%2F%2Fmatragut.wordpress.com%2F2010%2F03%2F18%2Fproblemas-de-genetica-2%2F
TEMA 3: GENÉTICA MENDELIANA
Cada ser vivo tiene unos rasgos, unas características que comparte con los de su especie, como son la forma y el aspecto externo, el modo de vida, el comportamiento, el tipo de alimentación, etc; cada ser vivo se parece a sus progenitores, pero siempre es diferente, hecho que ya observó el propio Darwin, pero, ¿qué es en realidad lo que nos dan nuestros padres para parecernos a ellos?; ¿por qué desarrollamos dos piernas, o seis patas, o cuatro alas?, ¿por qué huimos de un predador, o cazamos a una presa, o bailamos delante de una hembra?, ¿dónde reside la información que necesitamos para ser como somos?, ¿cómo le pasamos esa información a nuestros descendientes?.
Estas cuestiones son las que intentan ser respondidas por la Genética, aquella parte de la Biología que estudia los caracteres de los seres vivos y su herencia, que estudia, en definitiva, cómo a partir de una molécula, el ADN, se desarrollan los caracteres, y cómo, a través de la reproducción, esa molécula pasa de padres a hijos, y por tanto los hijos pueden desarrollar los mismos caracteres que los padres...........
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/index.htm
Estas cuestiones son las que intentan ser respondidas por la Genética, aquella parte de la Biología que estudia los caracteres de los seres vivos y su herencia, que estudia, en definitiva, cómo a partir de una molécula, el ADN, se desarrollan los caracteres, y cómo, a través de la reproducción, esa molécula pasa de padres a hijos, y por tanto los hijos pueden desarrollar los mismos caracteres que los padres...........
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/index.htm
jueves, 20 de mayo de 2010
Científicos en EE UU anuncian la creación de la primera vida artificial
"Craig Venter diseña una primera versión de célula y presagia un futuro Sillicon Valley del diseño de organismos vivos ............"
http://www.elpais.com/articulo/sociedad/Cientificos/EE/UU/anuncian/creacion/primera/vida/artificial/elpepusoc/20100520elpepusoc_9/Tes
http://www.elpais.com/articulo/sociedad/Cientificos/EE/UU/anuncian/creacion/primera/vida/artificial/elpepusoc/20100520elpepusoc_9/Tes
DICCIONARIO DE GENÉTICA
"El Instituto Nacional de Investigación del Genoma Humano (NHGRI) ha creado el Glosario hablado de términos de genética ........"
http://www.genome.gov/sglossary.cfm
http://www.genome.gov/sglossary.cfm
jueves, 13 de mayo de 2010

martes, 4 de mayo de 2010
TEMA 2: LA INFORMACIÓN GENÉTICA
1- LOS ÁCIDOS NUCLEICOS
2- LA REPLICACIÓN DEL ADN
3- EL CONCEPTO DE GEN
4- LAS MUTACIONES
5- LA EXPRESIÓN DE LA INFORMACIÓN GENÉTICA
6- BIOTECNOLOGÍA
7- LA INGIENERÍA GENÉTICA Y APLICACIONES
2- LA REPLICACIÓN DEL ADN
3- EL CONCEPTO DE GEN
4- LAS MUTACIONES
5- LA EXPRESIÓN DE LA INFORMACIÓN GENÉTICA
6- BIOTECNOLOGÍA
7- LA INGIENERÍA GENÉTICA Y APLICACIONES
LA DIVISIÓN CELULAR
"La división celular es, en realidad, un proceso doble. Estos dos procesos son:
- la división nuclear, o CARIOCINESIS
- la división citoplásmica, o CITOCINESIS
Ambos procesos pueden darse asociados, uno detrás del otro, o de forma independiente, primero uno, y algún tiempo después el otro.........."
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos5.htm
- la división nuclear, o CARIOCINESIS
- la división citoplásmica, o CITOCINESIS
Ambos procesos pueden darse asociados, uno detrás del otro, o de forma independiente, primero uno, y algún tiempo después el otro.........."
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos5.htm
LOS CROMOSOMAS
"Los cromosomas (término que significa "cuerpos coloreados", por la intensidad con la que fijan determinados colorantes al ser teñidos para poder observarlos al microscopio), son un componente del núcleo celular ............................"
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos4.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos4.htm
lunes, 22 de marzo de 2010
TEORÍA ENDOSIMBIÓTICA
La teoría endosimbiótica postula que algunos orgánulos propios de las células eucariotas, especialmente cloroplastos y mitocondrias, habrían tenido su origen en organismos procariotas que después de ser englobados por otro microorganismo habrían establecido una relación endosimbiótica con éste. Se especula con que las mitocondrias provendrían de protebacterias alfa y los cloroplastos de cianobacterias.
La teoría endosimbiótica fue popularizada por Lynn Margulis en 1967, con el nombre de endosimbiosis en serie. En su libro de 1981, Margulis sostiene que las células eucariotas se originaron como comunidades de entidades que obraban recíprocamente y que terminaron en la fusión de varios organismos. En la actualidad, se acepta que las mitocondrias y los cloroplastos de los eucariontes procedan de la endosimbiosis. Pero la idea de que una espiroqueta endosimbiótica se convirtiera en los flagelos y cilios de los eucariontes no ha recibido mucha aceptación, debido a que estos no muestran semejanzas ultraestructurales con los flagelos de los procariontes y carecen de ADN.
Antecedentes del origen simbiogenético de las eucariotas
En 1883, el biólogo alemán Andreas Schimper propuso que la capacidad fotosintética de las células vegetales podía proceder de cianobacterias aun presentes en la naturaleza y con iguales capacidades.
A principios del siglo XX Ivan Wallin (anatomista estadounidense) llegó a la misma conclusión, y en 1910 Kostantin S. Mereschovky presentó la hipótesis según la cual el origen de las células eucariotas se encontraba en la fusión de varias bacterias diferentes.
En 1918 el biólogo francés Paul Portier llegaría a la conclusión de que las mitocondrias de las eucariotas habrían sido en su día bacterias de vida libre, ahora confinadas en el interior de estas eucariotas; Wallin llegaría en 1925 a la misma conclusión.
Estos trabajos, minusvalorados en su tiempo, permanecieron olvidados hasta que Margulis, apoyándose en ellos, poniendo énfasis en las capacidades de las bacterias y la potencialidad de la simbiosis, formulara en 1967 la teoría endosimbiótica.
Teoria Endosimbiótica
La teoría endosimbiótica describe el paso de las células procariotas (células bacterianas, no nucleadas) a las células eucariotas (células nucleadas constituyentes de los procariontes y componentes de todos los pluricelulares) mediante incorporaciones simbiogenéticas sucesivas.
Margulis describe este paso en una serie de tres incorporaciones mediante las cuales, por la unión simbiogenética de bacterias, se originaron las células que conforman a los individuos de los otros cuatro reinos (protistas, animales, hongos y plantas).
Según la estimación más aceptada, hace 2.000 millones de años (aunque una horquilla posible podría descender a la cifra de 1.500 millones de años) la vida la componían multitud de bacterias diferentes, adaptadas a los diferentes medios. Margulis destacó también, la que debió ser una alta capacidad de adaptación de estas bacterias al cambiante e inestable ambiente de la Tierra en aquella época. Hoy se conocen más de veinte metabolismos diferentes usados por las bacterias frente al único usado por los pluricelulares: el aeróbico (que usan el oxígeno como fuente de energía; las plantas utilizan dos: aeróbico y fotosíntesis). Para Margulis, tal variedad revela las dificultades a las que las bacterias se tuvieron que enfrentar y su capacidad para aportar soluciones a esas dificultades.
Primera incorporación simbiogenética:
Una bacteria consumidora de azufre, que utilizaba el azufre y el calor como fuente de energía (arquea fermentadora o termoacidófila), se fusionó con una bacteria nadadora (espiroqueta) pasando a formar un nuevo organismo sumando sus características iníciales de forma sinérgica (en la que el resultado de la incorporación de dos o más unidades adquiere mayor valor que la suma de sus componentes). El resultado fue el primer eucarionte (unicelular eucariota) y ancestro único de todos los pluricelulares. El núcleo de la células de animales, plantas y hongos sería el resultado de la unión de estas dos bacterias.
A las características iniciales de ambas células se le sumó una nueva morfología más compleja con una nueva y llamativa resistencia al intercambio genético horizontal. El ADN quedó confinado en un núcleo interno separado del resto de la célula por una membrana.
Esta parte de la teoría (incorporación de la espiroqueta) no es aceptada en la actualidad, pues sólo la defienden Margulis y sus asociados. Ninguna de las homologías propuestas entre los flagelos de los eucariontes y de las espiroquetas ha resistido el escrutinio.
Segunda incorporación simbiogenética:
Este nuevo organismo todavía era anaeróbico, incapaz de metabolizar el oxígeno, ya que este gas suponía un veneno para él, por lo que viviría en medios donde este oxigeno, cada vez más presente, fuese escaso. En este punto, una nueva incorporación dotaría a este primigenio eucarionte de la capacidad para metabolizar oxigeno. Este nuevo endosombionte, originariamente bacteria respiradora de oxigeno de vida libre, se convertiría en las actuales mitocondrias presentes en las células eucariotas de los pluricelulares, posibilitando su éxito en un medio rico en oxígeno como ha llegado a convertirse el planeta Tierra. Los animales y hongos somos el resultado de esta segunda incorporación.

La teoría endosimbiótica fue popularizada por Lynn Margulis en 1967, con el nombre de endosimbiosis en serie. En su libro de 1981, Margulis sostiene que las células eucariotas se originaron como comunidades de entidades que obraban recíprocamente y que terminaron en la fusión de varios organismos. En la actualidad, se acepta que las mitocondrias y los cloroplastos de los eucariontes procedan de la endosimbiosis. Pero la idea de que una espiroqueta endosimbiótica se convirtiera en los flagelos y cilios de los eucariontes no ha recibido mucha aceptación, debido a que estos no muestran semejanzas ultraestructurales con los flagelos de los procariontes y carecen de ADN.
Antecedentes del origen simbiogenético de las eucariotas
En 1883, el biólogo alemán Andreas Schimper propuso que la capacidad fotosintética de las células vegetales podía proceder de cianobacterias aun presentes en la naturaleza y con iguales capacidades.
A principios del siglo XX Ivan Wallin (anatomista estadounidense) llegó a la misma conclusión, y en 1910 Kostantin S. Mereschovky presentó la hipótesis según la cual el origen de las células eucariotas se encontraba en la fusión de varias bacterias diferentes.
En 1918 el biólogo francés Paul Portier llegaría a la conclusión de que las mitocondrias de las eucariotas habrían sido en su día bacterias de vida libre, ahora confinadas en el interior de estas eucariotas; Wallin llegaría en 1925 a la misma conclusión.
Estos trabajos, minusvalorados en su tiempo, permanecieron olvidados hasta que Margulis, apoyándose en ellos, poniendo énfasis en las capacidades de las bacterias y la potencialidad de la simbiosis, formulara en 1967 la teoría endosimbiótica.
Teoria Endosimbiótica
La teoría endosimbiótica describe el paso de las células procariotas (células bacterianas, no nucleadas) a las células eucariotas (células nucleadas constituyentes de los procariontes y componentes de todos los pluricelulares) mediante incorporaciones simbiogenéticas sucesivas.
Margulis describe este paso en una serie de tres incorporaciones mediante las cuales, por la unión simbiogenética de bacterias, se originaron las células que conforman a los individuos de los otros cuatro reinos (protistas, animales, hongos y plantas).
Según la estimación más aceptada, hace 2.000 millones de años (aunque una horquilla posible podría descender a la cifra de 1.500 millones de años) la vida la componían multitud de bacterias diferentes, adaptadas a los diferentes medios. Margulis destacó también, la que debió ser una alta capacidad de adaptación de estas bacterias al cambiante e inestable ambiente de la Tierra en aquella época. Hoy se conocen más de veinte metabolismos diferentes usados por las bacterias frente al único usado por los pluricelulares: el aeróbico (que usan el oxígeno como fuente de energía; las plantas utilizan dos: aeróbico y fotosíntesis). Para Margulis, tal variedad revela las dificultades a las que las bacterias se tuvieron que enfrentar y su capacidad para aportar soluciones a esas dificultades.
Primera incorporación simbiogenética:
Una bacteria consumidora de azufre, que utilizaba el azufre y el calor como fuente de energía (arquea fermentadora o termoacidófila), se fusionó con una bacteria nadadora (espiroqueta) pasando a formar un nuevo organismo sumando sus características iníciales de forma sinérgica (en la que el resultado de la incorporación de dos o más unidades adquiere mayor valor que la suma de sus componentes). El resultado fue el primer eucarionte (unicelular eucariota) y ancestro único de todos los pluricelulares. El núcleo de la células de animales, plantas y hongos sería el resultado de la unión de estas dos bacterias.
A las características iniciales de ambas células se le sumó una nueva morfología más compleja con una nueva y llamativa resistencia al intercambio genético horizontal. El ADN quedó confinado en un núcleo interno separado del resto de la célula por una membrana.
Esta parte de la teoría (incorporación de la espiroqueta) no es aceptada en la actualidad, pues sólo la defienden Margulis y sus asociados. Ninguna de las homologías propuestas entre los flagelos de los eucariontes y de las espiroquetas ha resistido el escrutinio.
Segunda incorporación simbiogenética:
Este nuevo organismo todavía era anaeróbico, incapaz de metabolizar el oxígeno, ya que este gas suponía un veneno para él, por lo que viviría en medios donde este oxigeno, cada vez más presente, fuese escaso. En este punto, una nueva incorporación dotaría a este primigenio eucarionte de la capacidad para metabolizar oxigeno. Este nuevo endosombionte, originariamente bacteria respiradora de oxigeno de vida libre, se convertiría en las actuales mitocondrias presentes en las células eucariotas de los pluricelulares, posibilitando su éxito en un medio rico en oxígeno como ha llegado a convertirse el planeta Tierra. Los animales y hongos somos el resultado de esta segunda incorporación.

Tercera incorporación simbiogenética:
Esta tercera incorporación originó el Reino vegetal, las recientemente adquiridas células respiradoras de oxígeno fagocitarían bacterias fotosintéticas y algunas de ellas, haciéndose resistentes, pasarían a formar parte del organismo, los cloroplastos, originando a su vez un nuevo organismo capaz de sintetizar la energía procedente del Sol. Estos nuevos pluricelulares, las plantas, con su éxito, contribuyeron y contribuyen al éxito de animales y hongos.
En la actualidad permanecen las bacterias descendientes de aquellas que debieron, por incorporación, originar las células eucariotas; así como aquellos protistas que no participaron en alguna de las sucesivas incorporaciones.
No todos los biólogos aceptan la Teoría Simbiótica. El hecho de que el DNA en los cloroplastos y las mitocondrias, tendría que ocurrir algún procesó para transferir características hereditarias a lo que se convertiría en el núcleo. Algunos biólogos han ofrecido otra explicación para el desarrollo de células eucarióticas. Los primeros eucariotas formaron sus organelos por medio de invaginaciones y rompimientos de algunas regiones de la membrana celular.
Juan Esteban 12-11-2008
LA CÉLULA EUCARIOTA
ACTIVIDADES:
Crucigrama:
http://www.isftic.mepsyd.es/w3/eos/MaterialesEducativos/mem2009/celula_eucariota/alumno/actividades/eso3431.htm
Rallenar huecos:
http://www.isftic.mepsyd.es/w3/eos/MaterialesEducativos/mem2009/celula_eucariota/alumno/actividades/eso3432.htm
http://www.isftic.mepsyd.es/w3/eos/MaterialesEducativos/mem2009/celula_eucariota/alumno/actividades/eso3433.htm
LA CÉLULA PROCARIOTA
"Los organismos procariotas presentan una composición simple........"
http://www.ite.educacion.es/w3/recursos/bachillerato/bioygeo/ventana_hook/index.html
ACTIVIDADES
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/seruni-pluricelulares/actividad12.htm
http://www.ite.educacion.es/w3/recursos/bachillerato/bioygeo/ventana_hook/index.html
ACTIVIDADES
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/seruni-pluricelulares/actividad12.htm
jueves, 11 de marzo de 2010
LOS NIVELES DE ORGANIZACIÓN
En la materia viva existen varios grados de complejidad, denominados niveles de organización. Dentro de los mismos se pueden diferenciar niveles abióticos (materia no viva) y niveles bióticos ( materia viva, es decir con las tres funciones propias de los seres vivos). Los diferentes niveles serían:
1.- Nivel subatómico: integrado por las partículas subatómicas que forman los elementos químicos (protones, neutrones, electrones).
2.- Nivel atómico: son los átomos que forman los seres vivos y que denominamos bioelementos. Del total de elementos químicos del sistema periódico, aproximadamente un 70% de los mismos los podemos encontrar en la materia orgánica. Los elementos que ocupan cerca del 98% de todo el organismo son el carbono (C), el hidrógeno (H), el oxígeno (O), el nitrógeno (N), el fósforo (P) y el azufre (S). Alrededor del 2% está representado por el calcio (Ca), sodio (Na), Cloro (Cl), potasio (K) y magnesio (Mg). En una proporción menor al 0,1% están el hierro (Fe), yodo (I), zinc (Zn), cobre (Cu), y muchos otros.
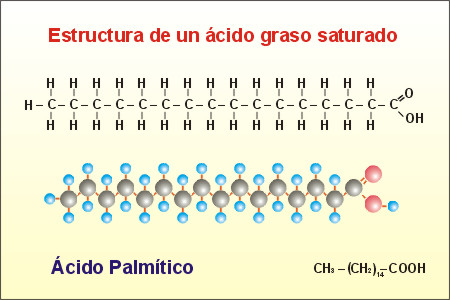
 3.- Nivel molecular: En él se incluyen las moléculas, formadas por la agrupación de átomos (bioelementos). A las moléculas orgánicas se les denomina Biomoléculas o Principios inmediatos. Estos Principios Inmediatos los podemos agrupar en dos categorías, inorgánicos (agua, sales minerales, iones, gases) y orgánicos (glúcidos, lípidos, proteínas y ácidos nucleicos).
3.- Nivel molecular: En él se incluyen las moléculas, formadas por la agrupación de átomos (bioelementos). A las moléculas orgánicas se les denomina Biomoléculas o Principios inmediatos. Estos Principios Inmediatos los podemos agrupar en dos categorías, inorgánicos (agua, sales minerales, iones, gases) y orgánicos (glúcidos, lípidos, proteínas y ácidos nucleicos).



En este nivel también debemos agrupar las macromoléculas y los virus. Las primeras resultan de la unión de monómeros (aminoácidos, nucleótidos, etc...) y los segundos son la unión de proteínas con ácidos nucleicos.
 4.- Nivel celular: donde nos encontramos a la célula (primer nivel con vida). Dos tipos de organizaciones celulares, Eucariota (células animales y vegetales) y Procariota (la bacteria). Los organismos unicelulares (Ej. Protozoos) viven con perfecta autonomía en el medio, pero en ocasiones nos podemos encontrar agrupaciones de células, las colonias, que no podemos considerar como seres pluricelulares por que a pesar de estar formados por miles de células cada una vive como un ser independiente.
4.- Nivel celular: donde nos encontramos a la célula (primer nivel con vida). Dos tipos de organizaciones celulares, Eucariota (células animales y vegetales) y Procariota (la bacteria). Los organismos unicelulares (Ej. Protozoos) viven con perfecta autonomía en el medio, pero en ocasiones nos podemos encontrar agrupaciones de células, las colonias, que no podemos considerar como seres pluricelulares por que a pesar de estar formados por miles de células cada una vive como un ser independiente.

5.- Nivel pluricelular: constituido por aquellos seres formados por más de una célula. Surge de la diferenciación y especialización celular. En él encontramos distintos niveles de complejidad: tejidos, órganos, sistemas y aparatos.
Mientras los tejidos son conjuntos de células de origen y forma parecida que realizan las mismas funciones, los órganos son un conjunto de tejidos diferentes que realizan actos concretos.
Los sistemas son conjuntos de órganos parecidos, al estar constituidos por los mismos tejidos, pero que realizan actos completamente independientes. Los aparatos (Ej. aparato digestivo), formados por órganos que pueden ser muy diferentes entre sí (Ej. dientes, lengua, estómago, etc...), realizan actos coordinados para constituir lo que se llama una función biológica (Ej. nutrición).
6.- Nivel de población: los individuos de la misma especie (aquellos que son capaces de reproducirse entre sí y tener descendencia fértil) se agrupan en poblaciones ( individuos de la misma especie que coinciden en el tiempo y en el espacio).
7.- Nivel de ecosistema: las poblaciones se asientan en una zona determinada donde se interrelacionan con otras poblaciones (COMUNIDAD O BIOCENOSIS) y con el medio no orgánico (Biotopo). Esta asociación configura el llamado ECOSISTEMA, objeto deestudio de los biólogos. Los ecosistemas son tan grande o tan pequeño como queramos, sin embargo el gran ecosistema terrestre lo forman la Biosfera (biocenosis) y el astro Tierra (biotopo).
1.- Nivel subatómico: integrado por las partículas subatómicas que forman los elementos químicos (protones, neutrones, electrones).
2.- Nivel atómico: son los átomos que forman los seres vivos y que denominamos bioelementos. Del total de elementos químicos del sistema periódico, aproximadamente un 70% de los mismos los podemos encontrar en la materia orgánica. Los elementos que ocupan cerca del 98% de todo el organismo son el carbono (C), el hidrógeno (H), el oxígeno (O), el nitrógeno (N), el fósforo (P) y el azufre (S). Alrededor del 2% está representado por el calcio (Ca), sodio (Na), Cloro (Cl), potasio (K) y magnesio (Mg). En una proporción menor al 0,1% están el hierro (Fe), yodo (I), zinc (Zn), cobre (Cu), y muchos otros.



Aminoácido


5.- Nivel pluricelular: constituido por aquellos seres formados por más de una célula. Surge de la diferenciación y especialización celular. En él encontramos distintos niveles de complejidad: tejidos, órganos, sistemas y aparatos.
Mientras los tejidos son conjuntos de células de origen y forma parecida que realizan las mismas funciones, los órganos son un conjunto de tejidos diferentes que realizan actos concretos.
Los sistemas son conjuntos de órganos parecidos, al estar constituidos por los mismos tejidos, pero que realizan actos completamente independientes. Los aparatos (Ej. aparato digestivo), formados por órganos que pueden ser muy diferentes entre sí (Ej. dientes, lengua, estómago, etc...), realizan actos coordinados para constituir lo que se llama una función biológica (Ej. nutrición).
6.- Nivel de población: los individuos de la misma especie (aquellos que son capaces de reproducirse entre sí y tener descendencia fértil) se agrupan en poblaciones ( individuos de la misma especie que coinciden en el tiempo y en el espacio).
7.- Nivel de ecosistema: las poblaciones se asientan en una zona determinada donde se interrelacionan con otras poblaciones (COMUNIDAD O BIOCENOSIS) y con el medio no orgánico (Biotopo). Esta asociación configura el llamado ECOSISTEMA, objeto deestudio de los biólogos. Los ecosistemas son tan grande o tan pequeño como queramos, sin embargo el gran ecosistema terrestre lo forman la Biosfera (biocenosis) y el astro Tierra (biotopo).
LA TEORÍA CELULAR
"Gracias al microscopio se conoce la estructura de los seres vivos. Por ello se sabe que todo ser vivo repite unas unidades estructurales que se llaman células. Todas las células cumplen las mismas funciones del ser vivo: autoconservación, autorregulación y autorreproducción....."
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/seruni-pluricelulares/contenidos3.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/seruni-pluricelulares/contenidos3.htm
TEMA 1: LA CÉLULA. UNIDAD DE VIDA
1- La Teoría Celular.
2- Los niveles de organización.
3- Tipos de organización celular.
Célula procariota y eucariota.
Célula animal y vegetal
4- El núcleo celular.
5- Los cromosomas.
6- El cariotipo: estudio e interpretación.
7- El ciclo celular.
Mitosis y meiosis.
2- Los niveles de organización.
3- Tipos de organización celular.
Célula procariota y eucariota.
Célula animal y vegetal
4- El núcleo celular.
5- Los cromosomas.
6- El cariotipo: estudio e interpretación.
7- El ciclo celular.
Mitosis y meiosis.
EXAMEN DE RECUPERACIÓN 2º TRIMESTRE
Lunes 15 de marzo de 2010
TEMA 9: Manifestaciones de la dinámica terrestre.
TEMA 7: El relieve y su modelado.
TEMA 10: La historia de nuestro planeta.
TEMA 4: Origen y evolución de los seres vivos.
TEMA 9: Manifestaciones de la dinámica terrestre.
TEMA 7: El relieve y su modelado.
TEMA 10: La historia de nuestro planeta.
TEMA 4: Origen y evolución de los seres vivos.
lunes, 8 de marzo de 2010
viernes, 5 de marzo de 2010
EL EQUILIBRIO PUNTUADO
Teoría del campo de la evolución biológica propuesta por Niles Eldredge y Stephen Jay Gould en 1972. Lo específico de la teoría del equilibrio puntuado tiene que ver con el tempo con el que las especies evolucionan. Según Eldredge y Gould, durante la mayor parte del tiempo de existencia de una especie ésta permanecería estable o con cambios menores (periodos de estasis), acumulándose cambio evolutivo durante el proceso de especiación (formación de una especie nueva), que sería una especie de revolución genética breve en términos geológicos. No se discute el carácter gradual del cambio evolutivo, sino que se niega la uniformidad de su ritmo.
Darwin era un estricto seguidor de este lema: “Natura non facit saltum”, la frase latina, atribuida a Linneo, mantiene que la naturaleza no da saltos. A pesar de que el registro fósil de la época no ofrecía apoyo alguno al cambio gradual. Darwin argumentaba que el registro fósil era imperfecto e incompleto: vemos los cambios abruptos porque nos faltan los pasos intermedios.
En las últimas décadas, Niles Eldredge y Stephen Jay Gould se han esforzado en argumentar que la teoría moderna de la evolución no tiene necesidad del gradualismo, y es éste el que se debe abandonar, no el darwinismo.
En su caso, la argumentación se refiere a la variación morfológica, y no a la molecular. Mientras que los neutralistas mantienen que el ritmo de evolución es más regular de lo que admite la teoría sintética, los puntualistas sostienen que el ritmo de evolución morfológica es menos regular de lo que esa hipótesis requiere. Los puntualistas niegan que el registro fósil sea incompleto. Sostienen que la aparición súbita de nuevas especies fósiles refleja que su formación se sigue a través de explosiones evolutivas, después de los cuales la especie sufrirá pocos cambios durante millones de años.
 Parte de esa aparente persistencia del registro puede ser consecuencia del fenómeno llamado evolución en mosaico, descrito por primera vez por Sir Gavin de Beer: el ritmo de cambio de las diferentes partes de un organismo no es uniforme en el transcurso de la evolución.
Parte de esa aparente persistencia del registro puede ser consecuencia del fenómeno llamado evolución en mosaico, descrito por primera vez por Sir Gavin de Beer: el ritmo de cambio de las diferentes partes de un organismo no es uniforme en el transcurso de la evolución.
La teoría del equilibrio puntuado no sólo se refiere al ritmo de la evolución, sino también a su curso. Eldredge y Gould postulan que la anagénesis (los cambios morfológicos experimentados por un mismo linaje) y la cladogénesis (la división de una especie en dos) están relacionadas causalmente. Mantienen que se da una breve aceleración del cambio morfológico precisamente cuando una población de censo reducido diverge de su especie original para formar otra nueva. La noción contraria, que los puntualistas atribuyen a la teoría sintética, consiste en que el cambio morfológico gradual lleva consigo su división en razas y subespecies mucho antes de que pueda afirmarse que han surgido especies nuevas.
En realidad, el cambio evolutivo sigue estos dos patrones, y otros muchos. El cambio morfológico y la aparición de mecanismos de aislamiento reproductor son fenómenos diferentes que pueden darse al mismo tiempo o por separado.
En palabras del propio Gould, en su libro de ensayos “El pulgar del panda”, responde a la pregunta de qué debería mostrar el registro fósil. Las especies deberían resultar estáticas en su territorio porque nuestros fósiles son los restos de grandes poblaciones centrales. En cualquier área local habitada por antecesores, una especie descendiente debería aparecer súbitamente por migración de la región periférica en la que evolucionó. En la propia zona periférica podríamos encontrar evidencias de la especiación, pero tan buena fortuna resultaría marcadamente infrecuente dada la velocidad a la que se produce el evento en una población tan pequeña. Así pues, el registro fósil es una fidedigna representación de lo que predice la teoría evolutiva, y no un pobre vestigio de lo que realmente haya ocurrido.
Eldredge y Gould llamaron a este modelo el de equilibrios puntuados. Las estirpes cambian poco durante la mayor parte de su historia, pero ocasionalmente esta tranquilidad se ve puntuada por rápidos procesos de especiación.
Gould también aclara en este mismo libro que no pretende mantener la verdad única del cambio puntuacional, sino ofrecer otras alternativas al cambio evolutivo. Como él dice, el gradualismo funciona bien en ocasiones.
Francisco Gil • Mayo 14, 2007
http://www.cienciaysociedad.info/
Darwin era un estricto seguidor de este lema: “Natura non facit saltum”, la frase latina, atribuida a Linneo, mantiene que la naturaleza no da saltos. A pesar de que el registro fósil de la época no ofrecía apoyo alguno al cambio gradual. Darwin argumentaba que el registro fósil era imperfecto e incompleto: vemos los cambios abruptos porque nos faltan los pasos intermedios.
En las últimas décadas, Niles Eldredge y Stephen Jay Gould se han esforzado en argumentar que la teoría moderna de la evolución no tiene necesidad del gradualismo, y es éste el que se debe abandonar, no el darwinismo.
En su caso, la argumentación se refiere a la variación morfológica, y no a la molecular. Mientras que los neutralistas mantienen que el ritmo de evolución es más regular de lo que admite la teoría sintética, los puntualistas sostienen que el ritmo de evolución morfológica es menos regular de lo que esa hipótesis requiere. Los puntualistas niegan que el registro fósil sea incompleto. Sostienen que la aparición súbita de nuevas especies fósiles refleja que su formación se sigue a través de explosiones evolutivas, después de los cuales la especie sufrirá pocos cambios durante millones de años.

La teoría del equilibrio puntuado no sólo se refiere al ritmo de la evolución, sino también a su curso. Eldredge y Gould postulan que la anagénesis (los cambios morfológicos experimentados por un mismo linaje) y la cladogénesis (la división de una especie en dos) están relacionadas causalmente. Mantienen que se da una breve aceleración del cambio morfológico precisamente cuando una población de censo reducido diverge de su especie original para formar otra nueva. La noción contraria, que los puntualistas atribuyen a la teoría sintética, consiste en que el cambio morfológico gradual lleva consigo su división en razas y subespecies mucho antes de que pueda afirmarse que han surgido especies nuevas.
En realidad, el cambio evolutivo sigue estos dos patrones, y otros muchos. El cambio morfológico y la aparición de mecanismos de aislamiento reproductor son fenómenos diferentes que pueden darse al mismo tiempo o por separado.
En palabras del propio Gould, en su libro de ensayos “El pulgar del panda”, responde a la pregunta de qué debería mostrar el registro fósil. Las especies deberían resultar estáticas en su territorio porque nuestros fósiles son los restos de grandes poblaciones centrales. En cualquier área local habitada por antecesores, una especie descendiente debería aparecer súbitamente por migración de la región periférica en la que evolucionó. En la propia zona periférica podríamos encontrar evidencias de la especiación, pero tan buena fortuna resultaría marcadamente infrecuente dada la velocidad a la que se produce el evento en una población tan pequeña. Así pues, el registro fósil es una fidedigna representación de lo que predice la teoría evolutiva, y no un pobre vestigio de lo que realmente haya ocurrido.
Eldredge y Gould llamaron a este modelo el de equilibrios puntuados. Las estirpes cambian poco durante la mayor parte de su historia, pero ocasionalmente esta tranquilidad se ve puntuada por rápidos procesos de especiación.
Gould también aclara en este mismo libro que no pretende mantener la verdad única del cambio puntuacional, sino ofrecer otras alternativas al cambio evolutivo. Como él dice, el gradualismo funciona bien en ocasiones.
Francisco Gil • Mayo 14, 2007
http://www.cienciaysociedad.info/
LAS ESPECIES Y LA ESPECIACIÓN
Especiación y Biodiversidad
Una entrevista original de ActionBioscience.org a Edward O. Wilson
- ¿Cómo se define una especie biológica?
- Wilson: El concepto de especie biológica es un concepto clásico. Se aplica, sin embargo, a especies que se reproducen sexualmente y se define de la siguiente manera: una especie es una población o serie de poblaciones de individuos que se cruzan libremente unos con los otros. En otras palabras, una especie es un elemento genético más o menos aislado, evolucionando por si mismo.
- Este concepto implica un pool genético aislado. ¿Es esto universal?
- El concepto de las especies no es universal.Wilson: No es universal. Puede ser aplicado a la gran mayoría de los animales y a una gran parte de las plantas que producen flores. Puede ser aplicable a muchos tipos de microorganismos. Pero hay muchos otros organismos, especialmente plantas y microorganismos, a los cuales no se les puede aplicar este concepto, o en el mejor de los casos, se les aplica con dificultad. Obviamente, no se puede aplicar si no hay reproducción sexual.
En el caso de las especies asexuales, es posible clasificarlas arbitrariamente basandose en diferencias genéticas. Por ejemplo, un estándar aplicado a bacterias es cuando dos cepas de ellas difieren tanto como un 30% en su ADN; en este caso se les puede tratar como dos especies diferentes. Por supuesto, esto es arbitrario, pero funciona razonablemente bien.
- ¿Cuales son los orígenes de la biodiversidad?
- Las especies tienden a evolucionar rápidamente en islas.Wilson: En primer lugar, es el ciclo aparente que siguen las especies al popular una nueva área y diversificarse. Cuando se forma una isla o un archipiélago, por ejemplo, o cuando la biodiversidad original de un área desaparece por una glaciación u otro evento físico mayor, ocurre una inundación de especies inmigrantes. Ellas interactúan y forman una comunidad que llamamos un ecosistema. Si la nueva área no se perturba, entonces típicamente ocurre un episodio de evolución rápida — una adaptación de nuevas especies al ambiente. Si existe suficiente espacio y partes geográficamente aisladas en esa área para sostener poblaciones que tienen poco contacto una con la otra, entonces también tenemos formación rápida de especies.
La especiación se desacelera después del primer estallido de diversificación.Un ejemplo típico sería Hawaii. La evidencia muestra que el archipiélago se llenó con un número pequeño de especies y que ellas evolucionaron y se diversificaron en muchas especies en un período de tiempo relativamente corto, con lo cual quiero decir, cientos o miles de años. Esto es un tiempo corto comparado a la evolución que se ve en otras partes del mundo.
La diversificación finalmente alcanza niveles donde la flora y la fauna se estabilizan, ocurriendo la coadaptación. Esto quiere decir que una especie es dependiente de otra, como por ejemplo una especie especializada en comerse a otra. En este punto, la especiación desacelera. Es similar al crecimiento de un organismo — rápido al principio con un ensamblaje de partes que interactúan y finalmente un nivel de madurez que puede ser sostenido por un período de tiempo largo.
El número de especies incrementa mientras más nos acercamos al ecuador.El otro grupo de principios de la biodiversidad tiene que ver con las cantidades, o con lo que determina las cantidades finales. Sabemos, por ejemplo, que los bosques húmedos tropicales tienen muchas más especies por unidad de área que la tundra o los bosques coníferos del hemisferio norte. El número de especies en un área definida, i.e., en una milla cuadrada o en cien millas cuadradas, aumenta a medida que uno se aproxima al ecuador. El número también aumenta si uno va de islas pequeñas a islas grandes, como en el Caribe............................
- ¿Qué determina este aumento de las especies?
- Wilson: Parece que hay tres factores, a los cuales me gusta referirme como ESA. E es por energía, S por estabilidad (stability en inglés) y A por área.
La energía, la estabilidad y el área influencian el número de especies.Mientras más energía se encuentra disponible para la comunidad de especies que evoluciona, mayor número de especies habrá. Esto se maximiza a medida que uno se mueve hacia el ecuador. Mientras más estable sea una región, como en una región de clima constante, se más especies porque tienen más tiempo de adaptarse y encajar juntas. Mientras más grande el área es, más grande y diversa es la población. Por ejemplo, Sudamérica tiene más especies que las Indias Occidentales.
El trabajo en equipo de estas tres fuerzas dirigentes parece ser responsable de la gran cantidad de variación de especies en el mundo.
- ¿Cuál es la forma más prevalente de especiación?
- El comportamiento parece jugar un papel mayor en la especiación que el aislamiento geográfico.La especiación puede suceder con relativa rapidez pero es difícil de observar.Wilson: Si la especiación simpátrica ocurre tanto entre insectos — el grupo de organismos más diverso del planeta — tal y como lo sugieren nuestros estudios, podría entonces ser la forma más prevalente de especiación. La especiación simpátrica es un fenómeno más difícil de estudiar que el de la especiación alopátrica, la cual es más fácil y más clara de observar. Sin embargo puede ser muy prevalente, pero no lo sabemos.
Simpátrica se refiere a organismos similares en proximidad cercana pero que no se entrecruzan porque tienen diferencias en comportamiento, a pesar de que en teoría se podrían cruzar. Alopátrica se refiere a organismos similares que no se entrecruzan, a pesar de que en teoría podrían, porque están geográficamente separados.
- ¿Cuán rápido pueden estos y otros mecanismos producir especies nuevas?
- Wilson: Instantáneamente. Bueno, en unas pocas generaciones. Primero que todo, en plantas existe un mecanismo — llamado poliploidía — que, en un solo paso, puede crear una cepa que no se puede cruzar con el grupo original del cual vino. De hecho, esto se llama especiación instantánea.
La especiación simpátrica puede suceder a veces en unos pocos pasos y producir especies nuevas en períodos cortos. Por ejemplo, en ciertos tipos de moscas de la fruta una cepa puede preferir aparearse en un tipo diferente de plantas, y esto puede ocurrir en varias mutaciones. Otra cepa puede llegar a reproducirse en una estación diferente y esto puede pasar a causa de varias mutaciones. O puede haber incompatibilidad entre dos cepas que difieren en uno o unos pocos genes. Esto puede ocurrir por mutación o recombinación de genes existentes en un período corto de tiempo.
Entonces, en teoría es posible crear nuevas especies en un período de varios años, y es probable que la formación de algunas especies ocurre así de rápido, pero es difícil de observar. No estamos seguros de cuan ampliamente distribuida es la especiación rápida.
E.O. Wilson, Ph.D., es Conservador Honorario de Entomología en el Museo de Zoología Comparativa de la Universidad de Harvard, así como Profesor Emeritus de Investigación de la Universidad Pellegrino. El Dr. Wilson, considerado internacionalmente como el decano de la biodiversidad, ha recibido de numerosos honores, incluyendo la Medalla de Oro del Fondo Mundial de la Naturaleza (1990) por su trabajo en el área de la conservación. Él es parte de la Junta de Directores de The Nature Conservancy y del American Museum of Natural History. Dos de sus libros han recibido el Premio Pulitzer; su libro más reciente se titula El Futuro de la Vida (2002).
Tipos de especiación:
"La clave de la especiación es la evolución de diferencias genéticas entre las especies incipientes. Para que un linaje se divida irreversiblemente, las dos especies incipientes deben tener diferencias genéticas que se expresen de alguna forma que haga que no se produzcan apareamientos entre ellas o que, de producirse, sean infructuosos. No hace falta que se trate de enormes diferencias genéticas, un pequeño cambio en el desarrollo cronológico, la localización o los rituales de apareamiento podrían ser suficientes. Pero, eso sí, es necesario que haya alguna diferencia. Este cambio podría producirse por selección natural o por deriva genética................................"
http://www.sesbe.org/evosite/evo101/VC1aModesSpeciation.shtml.html
Una entrevista original de ActionBioscience.org a Edward O. Wilson
- ¿Cómo se define una especie biológica?
- Wilson: El concepto de especie biológica es un concepto clásico. Se aplica, sin embargo, a especies que se reproducen sexualmente y se define de la siguiente manera: una especie es una población o serie de poblaciones de individuos que se cruzan libremente unos con los otros. En otras palabras, una especie es un elemento genético más o menos aislado, evolucionando por si mismo.
- Este concepto implica un pool genético aislado. ¿Es esto universal?
- El concepto de las especies no es universal.Wilson: No es universal. Puede ser aplicado a la gran mayoría de los animales y a una gran parte de las plantas que producen flores. Puede ser aplicable a muchos tipos de microorganismos. Pero hay muchos otros organismos, especialmente plantas y microorganismos, a los cuales no se les puede aplicar este concepto, o en el mejor de los casos, se les aplica con dificultad. Obviamente, no se puede aplicar si no hay reproducción sexual.
En el caso de las especies asexuales, es posible clasificarlas arbitrariamente basandose en diferencias genéticas. Por ejemplo, un estándar aplicado a bacterias es cuando dos cepas de ellas difieren tanto como un 30% en su ADN; en este caso se les puede tratar como dos especies diferentes. Por supuesto, esto es arbitrario, pero funciona razonablemente bien.
- ¿Cuales son los orígenes de la biodiversidad?
- Las especies tienden a evolucionar rápidamente en islas.Wilson: En primer lugar, es el ciclo aparente que siguen las especies al popular una nueva área y diversificarse. Cuando se forma una isla o un archipiélago, por ejemplo, o cuando la biodiversidad original de un área desaparece por una glaciación u otro evento físico mayor, ocurre una inundación de especies inmigrantes. Ellas interactúan y forman una comunidad que llamamos un ecosistema. Si la nueva área no se perturba, entonces típicamente ocurre un episodio de evolución rápida — una adaptación de nuevas especies al ambiente. Si existe suficiente espacio y partes geográficamente aisladas en esa área para sostener poblaciones que tienen poco contacto una con la otra, entonces también tenemos formación rápida de especies.
La especiación se desacelera después del primer estallido de diversificación.Un ejemplo típico sería Hawaii. La evidencia muestra que el archipiélago se llenó con un número pequeño de especies y que ellas evolucionaron y se diversificaron en muchas especies en un período de tiempo relativamente corto, con lo cual quiero decir, cientos o miles de años. Esto es un tiempo corto comparado a la evolución que se ve en otras partes del mundo.
La diversificación finalmente alcanza niveles donde la flora y la fauna se estabilizan, ocurriendo la coadaptación. Esto quiere decir que una especie es dependiente de otra, como por ejemplo una especie especializada en comerse a otra. En este punto, la especiación desacelera. Es similar al crecimiento de un organismo — rápido al principio con un ensamblaje de partes que interactúan y finalmente un nivel de madurez que puede ser sostenido por un período de tiempo largo.
El número de especies incrementa mientras más nos acercamos al ecuador.El otro grupo de principios de la biodiversidad tiene que ver con las cantidades, o con lo que determina las cantidades finales. Sabemos, por ejemplo, que los bosques húmedos tropicales tienen muchas más especies por unidad de área que la tundra o los bosques coníferos del hemisferio norte. El número de especies en un área definida, i.e., en una milla cuadrada o en cien millas cuadradas, aumenta a medida que uno se aproxima al ecuador. El número también aumenta si uno va de islas pequeñas a islas grandes, como en el Caribe............................
- ¿Qué determina este aumento de las especies?
- Wilson: Parece que hay tres factores, a los cuales me gusta referirme como ESA. E es por energía, S por estabilidad (stability en inglés) y A por área.
La energía, la estabilidad y el área influencian el número de especies.Mientras más energía se encuentra disponible para la comunidad de especies que evoluciona, mayor número de especies habrá. Esto se maximiza a medida que uno se mueve hacia el ecuador. Mientras más estable sea una región, como en una región de clima constante, se más especies porque tienen más tiempo de adaptarse y encajar juntas. Mientras más grande el área es, más grande y diversa es la población. Por ejemplo, Sudamérica tiene más especies que las Indias Occidentales.
El trabajo en equipo de estas tres fuerzas dirigentes parece ser responsable de la gran cantidad de variación de especies en el mundo.
- ¿Cuál es la forma más prevalente de especiación?
- El comportamiento parece jugar un papel mayor en la especiación que el aislamiento geográfico.La especiación puede suceder con relativa rapidez pero es difícil de observar.Wilson: Si la especiación simpátrica ocurre tanto entre insectos — el grupo de organismos más diverso del planeta — tal y como lo sugieren nuestros estudios, podría entonces ser la forma más prevalente de especiación. La especiación simpátrica es un fenómeno más difícil de estudiar que el de la especiación alopátrica, la cual es más fácil y más clara de observar. Sin embargo puede ser muy prevalente, pero no lo sabemos.
Simpátrica se refiere a organismos similares en proximidad cercana pero que no se entrecruzan porque tienen diferencias en comportamiento, a pesar de que en teoría se podrían cruzar. Alopátrica se refiere a organismos similares que no se entrecruzan, a pesar de que en teoría podrían, porque están geográficamente separados.
- ¿Cuán rápido pueden estos y otros mecanismos producir especies nuevas?
- Wilson: Instantáneamente. Bueno, en unas pocas generaciones. Primero que todo, en plantas existe un mecanismo — llamado poliploidía — que, en un solo paso, puede crear una cepa que no se puede cruzar con el grupo original del cual vino. De hecho, esto se llama especiación instantánea.
La especiación simpátrica puede suceder a veces en unos pocos pasos y producir especies nuevas en períodos cortos. Por ejemplo, en ciertos tipos de moscas de la fruta una cepa puede preferir aparearse en un tipo diferente de plantas, y esto puede ocurrir en varias mutaciones. Otra cepa puede llegar a reproducirse en una estación diferente y esto puede pasar a causa de varias mutaciones. O puede haber incompatibilidad entre dos cepas que difieren en uno o unos pocos genes. Esto puede ocurrir por mutación o recombinación de genes existentes en un período corto de tiempo.
Entonces, en teoría es posible crear nuevas especies en un período de varios años, y es probable que la formación de algunas especies ocurre así de rápido, pero es difícil de observar. No estamos seguros de cuan ampliamente distribuida es la especiación rápida.
E.O. Wilson, Ph.D., es Conservador Honorario de Entomología en el Museo de Zoología Comparativa de la Universidad de Harvard, así como Profesor Emeritus de Investigación de la Universidad Pellegrino. El Dr. Wilson, considerado internacionalmente como el decano de la biodiversidad, ha recibido de numerosos honores, incluyendo la Medalla de Oro del Fondo Mundial de la Naturaleza (1990) por su trabajo en el área de la conservación. Él es parte de la Junta de Directores de The Nature Conservancy y del American Museum of Natural History. Dos de sus libros han recibido el Premio Pulitzer; su libro más reciente se titula El Futuro de la Vida (2002).
Tipos de especiación:
"La clave de la especiación es la evolución de diferencias genéticas entre las especies incipientes. Para que un linaje se divida irreversiblemente, las dos especies incipientes deben tener diferencias genéticas que se expresen de alguna forma que haga que no se produzcan apareamientos entre ellas o que, de producirse, sean infructuosos. No hace falta que se trate de enormes diferencias genéticas, un pequeño cambio en el desarrollo cronológico, la localización o los rituales de apareamiento podrían ser suficientes. Pero, eso sí, es necesario que haya alguna diferencia. Este cambio podría producirse por selección natural o por deriva genética................................"
http://www.sesbe.org/evosite/evo101/VC1aModesSpeciation.shtml.html
jueves, 25 de febrero de 2010
PRUEBAS DE LA EVOLUCIÓN
"La evolución biológica es, posiblemente, el proceso más importante que afecta al conjunto de seres vivos que habitan en la Tierra, aunque este proceso no se de directamente sobre seres vivos determinados, ya que es un proceso que se prolonga mucho en el tiempo y tarda miles o millones de años en manifestarse; a pesar de ello, es un proceso imparable que comenzó con la aparición de la vida y desde entonces no ha perdido nada de vigor........."
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/5pruebas_de_la_evolucion.htm
ACTIVIDADES:
http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema6_4eso/Pruebas_evolucion.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/5pruebas_de_la_evolucion.htm
ACTIVIDADES:
http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema6_4eso/Pruebas_evolucion.htm
jueves, 18 de febrero de 2010
EL LAMARCKISMO
El lamarckismo fue una teoría propuesta en el siglo XIX por el biologo frances Jean-Baptiste Lamarck para explicar la evolució de las especies, también conocida como herencia de carácteres adquiridos, su formulación más simple postulaba que los individuos podían adquirir o mejorar carácteres físicos durante su vida y que estos eran transmitidos a su descendencia. De esta forma, las especies evolucionarían acumulando los carácteres útiles que habían adquirido en vida sus antepasados.
 "Así, para llegar a conocer las verdaderas causas de tantas formas diversas y de tantos hábitos diferentes como nos ofrecen los animales, es preciso considerar que las circunstancias infinitamente diversificadas, en las cuales se han encontrado los seres de cada raza, han producido para cada uno de ellos necesidades nuevas y cambios en sus hábitos necesariamente. Reconocida esta verdad, que nadie podrá negar, será fácil percibir cómo las nuevas necesidades han podido ser satisfechas y los nuevos hábitos adquiridos, si se presta alguna atención a las dos siguientes leyes de la Naturaleza, que siempre ha comprobado la observación: Primera ley: En todo animal que no ha traspasado el término de sus desarrollos, el uso frecuente y sostenido de un órgano cualquiera lo fortifica poco a poco, dándole una potencia proporcional a la duración de este uso, mientras que el desuso constante de tal órgano lo debilita y hasta le hace desaparecer.
"Así, para llegar a conocer las verdaderas causas de tantas formas diversas y de tantos hábitos diferentes como nos ofrecen los animales, es preciso considerar que las circunstancias infinitamente diversificadas, en las cuales se han encontrado los seres de cada raza, han producido para cada uno de ellos necesidades nuevas y cambios en sus hábitos necesariamente. Reconocida esta verdad, que nadie podrá negar, será fácil percibir cómo las nuevas necesidades han podido ser satisfechas y los nuevos hábitos adquiridos, si se presta alguna atención a las dos siguientes leyes de la Naturaleza, que siempre ha comprobado la observación: Primera ley: En todo animal que no ha traspasado el término de sus desarrollos, el uso frecuente y sostenido de un órgano cualquiera lo fortifica poco a poco, dándole una potencia proporcional a la duración de este uso, mientras que el desuso constante de tal órgano lo debilita y hasta le hace desaparecer.
Segunda ley: Todo lo que la Naturaleza hizo adquirir o perder a los individuos por la influencia de las circunstancias en que su raza se ha encontrado colocada durante largo tiempo, y consecuentemente por la influencia del empleo predominante de tal órgano, o por la de su desuso, la Naturaleza lo conserva por la generación en los nuevos individuos, con tal de que los cambios adquiridos sean comunes a los dos sexos, o a los que han producido estos nuevos individuos."
Lamarck, Filosofía zoológica, pp. 175-176.

Segunda ley: Todo lo que la Naturaleza hizo adquirir o perder a los individuos por la influencia de las circunstancias en que su raza se ha encontrado colocada durante largo tiempo, y consecuentemente por la influencia del empleo predominante de tal órgano, o por la de su desuso, la Naturaleza lo conserva por la generación en los nuevos individuos, con tal de que los cambios adquiridos sean comunes a los dos sexos, o a los que han producido estos nuevos individuos."
Lamarck, Filosofía zoológica, pp. 175-176.
LA DIVERSIDAD QUÍMICA DEL COSMOS EN UN METEORÍTO
"El famoso meteorito de Murchison, encontrado hace 40 años en Australia, contiene un impresionante arsenal de millones de componentes orgánicos, muchos más de los que se habían identificado hasta ahora, según han demostrado los últimos análisis de un grupo de químicos europeos.
El hallazgo pone de manifiesto que el Sistema Solar que acoge nuestro planeta contiene una variedad de química orgánica que sobrepasa la diversidad molecular que se encuentra en la Tierra. En concreto, se han localizado más de 14.000 diferentes elementos en la composición de la roca espacial.
El meteorito Murchison es uno de los tres más famosos localizados hasta ahora (los otros son el Allende, que cayó en México, y el Orgueil, en Francia) debido a su riqueza química. Cayó cerca de una localidad australiana, a la que debe su nombre, un 28 de septiembre de 1969. "Estos meteoritos son fundamentales para investigar la Tierra primitiva y entender las interacciones químicas y mineralógicas primigenias que contribuyeron al origen de la vida", explica el investigador del Centro de Astrobiología del CSIC-INTA Jesús Martínez Frías.
Los estudios que a lo largo de los años han hecho los investigadores en esta condrita (roca)carbonácea demostraron que contenía muchos aminoácidos fundamentales para la vida, si bien también dieron lugar a un intenso debate entre quienes defendían que se trataba de contaminación de origen terrestre y quienes aseguraban que habían venido del espacio exterior.

Posible contaminación
En este nuevo trabajo, publicado en Proceedings of National Academy of Science (PNAS), los autores, dirigidos por Philipe Schmitt-Kopplin, del Instituto alemán de Química Ecológica, han estudiado tres fragmentos del Murchison obtenidos en tres colecciones para diferenciar la posible contaminación del contenido original (estaban en Alaska, Japón y Luxemburgo).
Para analizarlos, utilizaron un espectrómetro de masa muy sofisticado que identificó 14.197 elementos en su composición, que podrían dar lugar a varios millones de estructuras moleculares. "Esta complejidad sugiere que la diversidad química extraterrestre es muy alta cuando se la compara con la bioquímica terrestre", aseguran los investigadores.
Además, han logrado determinar las interacciones que se han producido en el meteorito entre compuestos orgánicos (carbono, oxígeno, hidrógeno y nitrógeno) e inorgánicos (sulfuro de hierro y níquel).
Para Martínez Frías este hallazgo es especialmente importante porque "relaciona los meteoritos con las komatiitas, que son las lavas primitivas de la Tierra, las rocas más antiguas, y que también se han encontrado asociadas a sulfuros de hierro, lo que confirmaría su relación con los meteoritos que formaron nuestro planeta y que dieron lugar a la vida".
Se cree que el Murchison se formó en los primeros días del Sistema Solar, quizás antes que el Sol, y que atravesó las nubes de ese primitivo sistema recogiendo elementos orgánicos. Por ello, los autores sugieren que trazando la secuencia de las moléculas orgánicas se puede conocer la cronología de la formación y alteración de esas moléculas. En definitiva, un viaje al pasado para entender qué ocurrió."
Rosa M. Tristán
EL MUNDO
Madrid martes 16/02/2010 18:38 horas
El hallazgo pone de manifiesto que el Sistema Solar que acoge nuestro planeta contiene una variedad de química orgánica que sobrepasa la diversidad molecular que se encuentra en la Tierra. En concreto, se han localizado más de 14.000 diferentes elementos en la composición de la roca espacial.
El meteorito Murchison es uno de los tres más famosos localizados hasta ahora (los otros son el Allende, que cayó en México, y el Orgueil, en Francia) debido a su riqueza química. Cayó cerca de una localidad australiana, a la que debe su nombre, un 28 de septiembre de 1969. "Estos meteoritos son fundamentales para investigar la Tierra primitiva y entender las interacciones químicas y mineralógicas primigenias que contribuyeron al origen de la vida", explica el investigador del Centro de Astrobiología del CSIC-INTA Jesús Martínez Frías.
Los estudios que a lo largo de los años han hecho los investigadores en esta condrita (roca)carbonácea demostraron que contenía muchos aminoácidos fundamentales para la vida, si bien también dieron lugar a un intenso debate entre quienes defendían que se trataba de contaminación de origen terrestre y quienes aseguraban que habían venido del espacio exterior.

Posible contaminación
En este nuevo trabajo, publicado en Proceedings of National Academy of Science (PNAS), los autores, dirigidos por Philipe Schmitt-Kopplin, del Instituto alemán de Química Ecológica, han estudiado tres fragmentos del Murchison obtenidos en tres colecciones para diferenciar la posible contaminación del contenido original (estaban en Alaska, Japón y Luxemburgo).
Para analizarlos, utilizaron un espectrómetro de masa muy sofisticado que identificó 14.197 elementos en su composición, que podrían dar lugar a varios millones de estructuras moleculares. "Esta complejidad sugiere que la diversidad química extraterrestre es muy alta cuando se la compara con la bioquímica terrestre", aseguran los investigadores.
Además, han logrado determinar las interacciones que se han producido en el meteorito entre compuestos orgánicos (carbono, oxígeno, hidrógeno y nitrógeno) e inorgánicos (sulfuro de hierro y níquel).
Para Martínez Frías este hallazgo es especialmente importante porque "relaciona los meteoritos con las komatiitas, que son las lavas primitivas de la Tierra, las rocas más antiguas, y que también se han encontrado asociadas a sulfuros de hierro, lo que confirmaría su relación con los meteoritos que formaron nuestro planeta y que dieron lugar a la vida".
Se cree que el Murchison se formó en los primeros días del Sistema Solar, quizás antes que el Sol, y que atravesó las nubes de ese primitivo sistema recogiendo elementos orgánicos. Por ello, los autores sugieren que trazando la secuencia de las moléculas orgánicas se puede conocer la cronología de la formación y alteración de esas moléculas. En definitiva, un viaje al pasado para entender qué ocurrió."
Rosa M. Tristán
EL MUNDO
Madrid martes 16/02/2010 18:38 horas
lunes, 15 de febrero de 2010
FIJISMO Y EVOLUCIONISMO
"Una vez que la vida surge sobre la Tierra, se nos plantea un nuevo interrogante: ¿cómo a partir de una sola célula han podido aparecer todas las especies tan diferentes que existen hoy día?..........."
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/3evolucion_de_los_seres_vivos.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/3evolucion_de_los_seres_vivos.htm
REVIVIENDO BACTERIAS Y LA TEORIA DE LA PANSPERMIA
"Científicos norteamericanos han conseguido revivir bacterias congeladas procedentes de la Antártida, con edades de entre un millón y ocho millones de años.
La única diferencia entre las bacterias más antiguas y las mas modernas es que las que tenían ocho millones de años tenían el ADN bastante dañado, con lo cual ya no eran viables.
Esto ha servido para determinar que la teoría de la Panspermia puede tener cierta base, o por lo menos que la vida podría viajar de un planeta a otro a bordo de cometas, siempre y cuando el viaje no dure más de 8 millones de años, ya que los microorganismos llegarían a su destinos con el ADN totalmente deteriorado.”
Guillermo Carvajal
Más sobre panspermia:
http://www.panspermia.org/
La única diferencia entre las bacterias más antiguas y las mas modernas es que las que tenían ocho millones de años tenían el ADN bastante dañado, con lo cual ya no eran viables.
Esto ha servido para determinar que la teoría de la Panspermia puede tener cierta base, o por lo menos que la vida podría viajar de un planeta a otro a bordo de cometas, siempre y cuando el viaje no dure más de 8 millones de años, ya que los microorganismos llegarían a su destinos con el ADN totalmente deteriorado.”
Guillermo Carvajal
Más sobre panspermia:
http://www.panspermia.org/
LAS PRINCIPALES HIPÓTESIS SOBRE EL ORIGEN DE LA VIDA
HIPÓTESIS DE LA SISTESIS PREBIÓTICA
"Hoy en día la teoría aceptada para explicar el origen de la vida es la que se basa en la hipótesis química expuesta por el ruso A. Oparin y el inglés Haldane en 1923......................"
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/2origen_quimico_vida.htm
HIPÓTEISIS DE LA PANSPERMIA
Es la hipótesis que sugiere que las “semillas” o la esencia de la vida prevalecen diseminadas por todo el universo y que la vida comenzó en la Tierra gracias a la llegada de tales semillas a nuestro planeta.
Estas ideas tienen su origen en algunas de las consideraciones del filósofo griego Anaxágoras. El término acuñado por el biólogo alemán Hermann Ritcher en 1865 usando el griego παν- (pan = todo) y σπερμα (sperma = semilla).
Fue en 1908 cuando el químico sueco Svante August Arrhenius usó la palabra panspermia para explicar el comienzo de la vida en la tierra. El astrónomo Fred Hoyle también apoyó la idea de la panspermia.
Existen pruebas de bacterias capaces de sobrevivir largos períodos de tiempo incluso en el espacio exterior, lo que apoyaría el mecanismo subyacente de este proceso. Estudios recientes en la India apoyan la hipótesis.[Otros han hallado bacterias en la atmósfera a altitudes de más de 40 km donde, aunque no se espera que se produzcan mezclas con capas inferiores, pueden haber llegado desde éstas.
Bacterias Streptococcus mitis que fueron llevadas a la Luna por accidente en la Surveyor 3 en 1967, pudieron ser revividas sin dificultad cuando llegaron de vuelta a la Tierra tres años después.
Una posible consecuencia de la panspermia sería que la vida en todo el Universo poseería una base bioquímica similar, a menos que hubiera más de una fuente original de vida. El mayor inconveniente de esta teoría es que no resuelve el problema inicial de cómo surgió la vida, si no que se limita a mover la responsabilidad del origen a otro lugar. Otra objeción a la panspermia es que las bacterias no sobrevivirían a las altísimas temperaturas y las fuerzas involucradas en un impacto contra la Tierra, aunque no se ha llegado aún a posiciones concluyentes en este punto (ni a favor ni en contra), pues se conocen algunas especies de bacterias extremófilas capaces de soportar condiciones de radiación, temperatura y presión extremas que hacen pensar que la vida pudiera adquirir formas insospechadamente resistentes.
El análisis del meteorito ALH 84001, generalmente considerado como originado en el planeta Marte, sugiere que contiene estructuras que podrían haber sido causadas por formas de vida microscópica. Ésta es hasta la fecha la única indicación de vida extraterrestre y aún es muy controvertida.
Por otro lado, existe el meteorito Murchinson, que contiene uracilo y xantina, dos precursores de las moléculas que configuran el ARN y el ADN. (Wikipedia)
En la actualidad, la teoría de panspermia tiene tres variantes a saber:
Panspermia Planetaria: Teoría que sugiere que la vida en la Tierra se origino en otro cuerpo del sistema solar, como Marte, y llego a nuestro planeta en el interior de un meteorito.
Panspermia Estelar: Teoría que sugiere que la vida en la Tierra se origino más allá del sistema solar, en las nebulosas o en otros sistemas planetarios y llego a nuestro planeta en el interior de cometas procedentes de esos distantes lugares del Universo.
Panspermia Dirigida: Teoría que sugiere que la vida en la Tierra se origino a causa de microorganismos enviados deliberadamente en el interior de cápsulas espaciales por civilizaciones extraterrestres con el objetivo de sembrar la vida por el Universo.
"Hoy en día la teoría aceptada para explicar el origen de la vida es la que se basa en la hipótesis química expuesta por el ruso A. Oparin y el inglés Haldane en 1923......................"
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/2origen_quimico_vida.htm
HIPÓTEISIS DE LA PANSPERMIA
Es la hipótesis que sugiere que las “semillas” o la esencia de la vida prevalecen diseminadas por todo el universo y que la vida comenzó en la Tierra gracias a la llegada de tales semillas a nuestro planeta.
Estas ideas tienen su origen en algunas de las consideraciones del filósofo griego Anaxágoras. El término acuñado por el biólogo alemán Hermann Ritcher en 1865 usando el griego παν- (pan = todo) y σπερμα (sperma = semilla).
Fue en 1908 cuando el químico sueco Svante August Arrhenius usó la palabra panspermia para explicar el comienzo de la vida en la tierra. El astrónomo Fred Hoyle también apoyó la idea de la panspermia.
Existen pruebas de bacterias capaces de sobrevivir largos períodos de tiempo incluso en el espacio exterior, lo que apoyaría el mecanismo subyacente de este proceso. Estudios recientes en la India apoyan la hipótesis.[Otros han hallado bacterias en la atmósfera a altitudes de más de 40 km donde, aunque no se espera que se produzcan mezclas con capas inferiores, pueden haber llegado desde éstas.
Bacterias Streptococcus mitis que fueron llevadas a la Luna por accidente en la Surveyor 3 en 1967, pudieron ser revividas sin dificultad cuando llegaron de vuelta a la Tierra tres años después.
Una posible consecuencia de la panspermia sería que la vida en todo el Universo poseería una base bioquímica similar, a menos que hubiera más de una fuente original de vida. El mayor inconveniente de esta teoría es que no resuelve el problema inicial de cómo surgió la vida, si no que se limita a mover la responsabilidad del origen a otro lugar. Otra objeción a la panspermia es que las bacterias no sobrevivirían a las altísimas temperaturas y las fuerzas involucradas en un impacto contra la Tierra, aunque no se ha llegado aún a posiciones concluyentes en este punto (ni a favor ni en contra), pues se conocen algunas especies de bacterias extremófilas capaces de soportar condiciones de radiación, temperatura y presión extremas que hacen pensar que la vida pudiera adquirir formas insospechadamente resistentes.
El análisis del meteorito ALH 84001, generalmente considerado como originado en el planeta Marte, sugiere que contiene estructuras que podrían haber sido causadas por formas de vida microscópica. Ésta es hasta la fecha la única indicación de vida extraterrestre y aún es muy controvertida.
Por otro lado, existe el meteorito Murchinson, que contiene uracilo y xantina, dos precursores de las moléculas que configuran el ARN y el ADN. (Wikipedia)
En la actualidad, la teoría de panspermia tiene tres variantes a saber:
Panspermia Planetaria: Teoría que sugiere que la vida en la Tierra se origino en otro cuerpo del sistema solar, como Marte, y llego a nuestro planeta en el interior de un meteorito.
Panspermia Estelar: Teoría que sugiere que la vida en la Tierra se origino más allá del sistema solar, en las nebulosas o en otros sistemas planetarios y llego a nuestro planeta en el interior de cometas procedentes de esos distantes lugares del Universo.
Panspermia Dirigida: Teoría que sugiere que la vida en la Tierra se origino a causa de microorganismos enviados deliberadamente en el interior de cápsulas espaciales por civilizaciones extraterrestres con el objetivo de sembrar la vida por el Universo.
EL ORIGEN DE LA VIDA
"Hasta el momento actual la ciencia no ha sido capaz de dar una explicación sobre lo que es la vida, aparte de estudiar sus características y sus manifestaciones. Además de explicar lo que es la vida, ha habido otro problema que ha preocupado al hombre desde siempre, y es el origen de la vida, ¿de dónde viene?, ¿cómo se ha formado?. Para explicar esto han existido dos grandes corrientes de pensamiento, la generación espontánea, idea que perduró hasta........"
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/1origen_de_la_vida.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/evolucion/1origen_de_la_vida.htm
jueves, 11 de febrero de 2010
TEMA 4: ORIGEN Y EVOLUCIÓN DE LOS SERES VIVOS
1- El origen de la vida
2- Principales hipótesis sobre el origen de la vida
3- La evolución biológica: Fijismo y evolucionismo
Fijismo
4- El lamarckismo
5- La teoría de la evolución de las especies por selección natural de Darwin
6- El origen de la variavilidad
7- La presión de selección y la adaptación
8- Pruebas de la evolución
9- El neodarwinismo
10- El equilibrio puntuado
11- Las especies y la especiación
12- El origen y la evolución humana
2- Principales hipótesis sobre el origen de la vida
3- La evolución biológica: Fijismo y evolucionismo
Fijismo
4- El lamarckismo
5- La teoría de la evolución de las especies por selección natural de Darwin
6- El origen de la variavilidad
7- La presión de selección y la adaptación
8- Pruebas de la evolución
9- El neodarwinismo
10- El equilibrio puntuado
11- Las especies y la especiación
12- El origen y la evolución humana
viernes, 5 de febrero de 2010


IDENTIFICACIÓN DE FÓSILES
"Un fósil es el resto de un organismo vivo que se ha conservado en el registro sedimentario mediante un proceso de fosilización. La fosilización es un proceso de mineralización mediante el cual los tejidos orgánicos son reemplazados por compuestos químicos nuevos. Generalmente, sólo se conservan las partes orgánicas que ya estaban mineralizadas (conchas, esqueletos, etc.). No obstante, cuando las condiciones del medio de sedimentación eran muy especiales (p.ej. baja energía, ambiente reductor, rápido enterramiento, etc.) se pueden conservar los tejidos blandos. Los icnofósiles son las señales de la actividad de un organismo que ha quedado conservadas en el registro geológico (p.ej. huellas de pisadas).
Los fósiles tienen un valor intrínseco paleobiológico, pues permiten estudiar múltiples aspectos biológicos, como por ejemplo, la evolución, la paleoecología, etc. Además, en los estudios de geología permiten realizar una serie de precisiones cronoestratigráficas y sedimentológicas importantes, pues ayudan, entre otras cosas, a determinar la edad relativa de las rocas, a correlacionar estratos, y aportan datos para establecer en detalle el ambiente de sedimentación..............."
http://www.uam.es/personal_pdi/ciencias/casado/GEORED/Fosiles/prac4.htm
Los fósiles tienen un valor intrínseco paleobiológico, pues permiten estudiar múltiples aspectos biológicos, como por ejemplo, la evolución, la paleoecología, etc. Además, en los estudios de geología permiten realizar una serie de precisiones cronoestratigráficas y sedimentológicas importantes, pues ayudan, entre otras cosas, a determinar la edad relativa de las rocas, a correlacionar estratos, y aportan datos para establecer en detalle el ambiente de sedimentación..............."
http://www.uam.es/personal_pdi/ciencias/casado/GEORED/Fosiles/prac4.htm
GEOCRONOLOGÍA RELATIVA
"Cuándo sucedió un hecho?; ¿cuándo se inició un proceso?; ¿qué sucedió antes?. Son preguntas importantes de contestar si queremos encontrar una explicación a la historia de la Tierra. Para ello no nos sirve entender el tiempo basándonos en la percepción humana, que es muy corta. Debemos buscar un concepto de tiempo que se adapte a la edad de la propia Tierra. A este concepto lo denominamos TIEMPO GEOLÓGICO.
a.- Cronología relativa y cronología absoluta
Cuando paseamos por una playa, vamos dejando huellas sobre las rizaduras de la arena. Si alguien pasa detrás de nosotros sabrá que primero se hicieron las rizaduras y luego las huellas; desconocerá en qué momento exacto sucedieron ambos hechos, pero sí sabrá cuál sucedió antes y cuál después. Es lo que llamamos CRONOLOGÍA RELATIVA.............."
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/contenidos2.htm
a.- Cronología relativa y cronología absoluta
Cuando paseamos por una playa, vamos dejando huellas sobre las rizaduras de la arena. Si alguien pasa detrás de nosotros sabrá que primero se hicieron las rizaduras y luego las huellas; desconocerá en qué momento exacto sucedieron ambos hechos, pero sí sabrá cuál sucedió antes y cuál después. Es lo que llamamos CRONOLOGÍA RELATIVA.............."
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/contenidos2.htm
martes, 2 de febrero de 2010
INTERPRETACIÓN DE CORTES GEOLÓGICOS
Página de la Universidad Autónoma de Madrid en la que se muestran como se interpretan cortes geológicos y algunos ejercícios para practicar.
http://www.uam.es/personal_pdi/ciencias/casado/GEORED/Cortes/test1.htm
Más ejercicios prácticos de interpretación de cortes geológicos:
http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema3_4eso/Interpretacion_corte7.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/actividad5.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/actividad6.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/historia_geologica/historia_geol_1.htm
http://www.uam.es/personal_pdi/ciencias/casado/GEORED/Cortes/test1.htm
Más ejercicios prácticos de interpretación de cortes geológicos:
http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema3_4eso/Interpretacion_corte7.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/actividad5.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/actividad6.htm
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/historia_geologica/historia_geol_1.htm
EDAD GEOCRONOLOGÍA ABSOLUTA
Página bastante interesante para profundizar en el método radiométrico para calcular la edad de las rocas:
http://ansatte.uit.no/kku000/webgeology/webgeology_files/spanish/edad-abs_8new.html
Actividades en las que tenemos que aplicar diversos métodos para calcular la edad absoluta de la Tierra.
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/tierra_cambia/actividad4.htm
martes, 26 de enero de 2010
TEMA 10: LA HISTORIA DE NUESTRO PLANETA
1- EL TIEMPO GEOLÓGICO Y LA EDAD DE LA TIERRA: UNIFORMITARISMO Y ACTUALISMO
2- GEOCRONOLOGÍA ABSOLUTA: EL MÉTODO RADIOMÉTRICO
3- GEOCRONOLOGÍA RELATIVA: PRINCIPIOS
-INTERPRETACIÓN DE CORTES GEOLÓGICOS
4- LOS FÓSILES Y SU APLICACIONES EN GEOCRONOLOGÍA
-IDENTIFICACIÓN DE FÓSILES Y SU APLICACIÓN
5- LA ESCALA DEL TIEMPO GEOLÓGICO
6- LA HISTORIA DE LA TIERRA
2- GEOCRONOLOGÍA ABSOLUTA: EL MÉTODO RADIOMÉTRICO
3- GEOCRONOLOGÍA RELATIVA: PRINCIPIOS
-INTERPRETACIÓN DE CORTES GEOLÓGICOS
4- LOS FÓSILES Y SU APLICACIONES EN GEOCRONOLOGÍA
-IDENTIFICACIÓN DE FÓSILES Y SU APLICACIÓN
5- LA ESCALA DEL TIEMPO GEOLÓGICO
6- LA HISTORIA DE LA TIERRA
martes, 19 de enero de 2010
SISTEMAS MORFOCLIMÁTICOS
"Aunque la litología y la estructura de las rocas condicionan el modelado, es el clima y, por tanto, los factores climáticos, lo que más influye en el relieve de una región. En consecuencia, pueden aparecer formas de relieve similares en lugares geográficos distantes entre ellos si tienen el mismo clima....."
http://www.kalipedia.com/ciencias-tierra-universo/tema/modelado-relieve/sistemas-morfoclimaticos.html?x=20070417klpcnatun_152.Kes
http://www.kalipedia.com/ciencias-tierra-universo/tema/modelado-relieve/sistemas-morfoclimaticos.html?x=20070417klpcnatun_152.Kes
FACTORES CONDICIONANTES DEL MODELADO
"El clima es el factor más influyente en el modelado del paisaje. No obstante, no es el único. Además del clima, los factores litológicos, estructurales, dinámicos, climáticos y antrópicos, combinados entre sí, dan lugar al relieve......"
http://www.kalipedia.com/ciencias-tierra-universo/tema/modelado-relieve/factores-condicionantes-modelado-relieve.html?x=20070417klpcnatun_150.Kes
http://www.kalipedia.com/popup/popupWindow.html?tipo=fotogaleria&url=20070417klpcnatun_67.Zes&seccion=Materias&subseccion=CienciasNaturales&subsubseccion=CienciasdelaTierraydelUniverso&tipo_contenido=COL&popw=804&poph=588
Ejemplo de la influencia que tienen las estructuras geológicas en el paisaje: oberva esta fotografía de satélite del Estado de Pennsylvania (Noreste de EEUU) y fijate en las franjas de color verde más intenso que representan zonas de bosques y montañas. Estas corresponderían a zonas donde los materiales estan plegados, este plegamiento junto con el tipo de roca que lo forman condiciona el relieve de la zona y le da el aspecto característico.
Ver mapa más grande
http://www.kalipedia.com/ciencias-tierra-universo/tema/modelado-relieve/factores-condicionantes-modelado-relieve.html?x=20070417klpcnatun_150.Kes
http://www.kalipedia.com/popup/popupWindow.html?tipo=fotogaleria&url=20070417klpcnatun_67.Zes&seccion=Materias&subseccion=CienciasNaturales&subsubseccion=CienciasdelaTierraydelUniverso&tipo_contenido=COL&popw=804&poph=588
Ejemplo de la influencia que tienen las estructuras geológicas en el paisaje: oberva esta fotografía de satélite del Estado de Pennsylvania (Noreste de EEUU) y fijate en las franjas de color verde más intenso que representan zonas de bosques y montañas. Estas corresponderían a zonas donde los materiales estan plegados, este plegamiento junto con el tipo de roca que lo forman condiciona el relieve de la zona y le da el aspecto característico.
Ver mapa más grande
jueves, 14 de enero de 2010
MODELADO CÁRSTICO
"Recibe este nombre el conjunto de acciones y procesos de modelado condicionados por la presencia de rocas carbonatadas, fundamentalmente calizas, que, siendo solubles bajo determinadas condiciones, dan lugar a morfologías y paisajes peculiares.........."
http://platea.pntic.mec.es/~cmarti3/GEO/mod/karst.htm


http://platea.pntic.mec.es/~cmarti3/GEO/mod/karst.htm

Travertinos

Actividades: http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema2_4eso/karst2.htm
MODELADO GLACIAL
"Se puede definir un glaciar como una masa de hielo policristalino procedente de nieve compactada y recristalizada. Esta masa de hielo se mueve desplazándose a favor de pendiente en un proceso de descarga desde la zona de acumulación hacia zonas bajas y/o marginales, en las que se produce la pérdida de hielo, ya sea por ablación (fusión, evaporación o sublimación) o por desmembramiento de la masa de hielo sobre aguas continentales o marinas........"
http://platea.pntic.mec.es/~cmarti3/GEO/mod/glac.htm
Actividades:
http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema2_4eso/glaciar_cards.htm
http://platea.pntic.mec.es/~cmarti3/GEO/mod/glac.htm
Actividades:
http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema2_4eso/glaciar_cards.htm
MODELADO LITORAL
"La acción erosiva del mar sobre las rocas del litoral se debe fundamentalmente a mecanismos relacionados con el efecto del oleaje: la presión ejercida por el agua y por el aire situado en grietas o fisuras, que es comprimido como por un émbolo al golpear la ola, más el impacto del material fragmentario que la ola arroja contra las rocas. Tampoco hay que olvidar la acción de la disolución que ejerce el agua y, en fin, la amplitud que a todas esas acciones proporciona la oscilación de las mareas......."
http://platea.pntic.mec.es/~cmarti3/GEO/mod/mar.htm

http://platea.pntic.mec.es/~cmarti3/GEO/mod/mar.htm
Retroceso de un acantilado

Actividades: http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema2_4eso/costas_cards.htm
MODELADO EÓLICO
"El desierto pedregoso o REG es el paisaje más extendido en las regiones áridas. Los relieves rocosos son atacados por la meteorización, fundamentalmente de tipo mecánico como consecuencia de la falta de agua, por termoclastia y crecimiento de cristales de sales. El viento terminará arrastrando las partículas más finas (esta acción es la deflación) y quedarán los cantos........"
http://platea.pntic.mec.es/~cmarti3/GEO/mod/arido.htm

http://platea.pntic.mec.es/~cmarti3/GEO/mod/arido.htm

Actividades: http://www.educa.madrid.org/web/cc.nsdelasabiduria.madrid/Ejercicios/Tema2_4eso/aridos1.htm
MODELADO TORRENCIAL
"Las aguas salvajes son aguas superficiales carentes de cauce y caudal fijos. Corresponden por tanto a fenómenos episódicos de precipitaciones más o menos intensas o fenómenos de deshielo repentino de origen climático o volcánico, por ejemplo. Su efecto erosivo puede llegar a ser muy importante, arrastrando grandes cantidades de materiales, destruyendo el suelo edáfico e, incluso, ocasionando avenidas y desencadenando movimientos de ladera. En todo caso, depende de factores como: cubierta vegetal del terreno, pendiente, tipo de material.
Los torrentes se encauzan en canales persistentes aunque el caudal es también temporal......."
http://platea.pntic.mec.es/~cmarti3/GEO/mod/salva.htm
Los torrentes se encauzan en canales persistentes aunque el caudal es también temporal......."
http://platea.pntic.mec.es/~cmarti3/GEO/mod/salva.htm
EL MODELADO FLUVIAL
"Los ríos se caracterizan por poseer un cauce más o menos fijo y un caudal permanente. Esto último es posible gracias a la aportación procedente de aguas subterráneas. Sin embargo,..........."
http://platea.pntic.mec.es/~cmarti3/GEO/mod/rios.htm
http://www.cienciasnaturales.es/FLUVIAL.swf
Cómo se forman las terrazas
http://platea.pntic.mec.es/~cmarti3/GEO/mod/fterraz.htm

http://platea.pntic.mec.es/~cmarti3/GEO/mod/rios.htm
http://www.cienciasnaturales.es/FLUVIAL.swf
Cómo se forman las terrazas
http://platea.pntic.mec.es/~cmarti3/GEO/mod/fterraz.htm

Actividades:
CUENCAS SEDIMENTARIAS Y AMBIENTES SEDIMENTARIOS
Las cuencas sedimentarias son depresiones naturales de la corteza terrestre, que pueden presentar dimensiones de varios miles de kilómetros cuadrados y en las que se acumulan materiales erosionados procedentes de otros lugares y transportados hasta allí por la acción del agua, el viento u otros medios. La cuenca sedimentaria por tanto delimita una zona geográfica en la que se produce hundimiento y por tanto acumulación de sedimentos. Las condiciones físicas (temperatura, etc..), químicas (concentración de sales, presencia de oxígeno, etc) y biológicas (vegetación, etc) en esa zona (normalmente muy amplia) varían de unos lugares a otros y a lo largo del tiempo. Estas condiciones físicas, químicas y biológicas es lo que se denominan ambientes sedimentarios, y van a condicionar el tipo de sedimentos y las estracturas sedimentarias que se van a producir. (ambientes sedimentarios: el conjunto de procesos físicos, químicos y biológicos que se encuentran afectando la sedimentación (Fraser, 1989))

Actividad:
http://recursos.cnice.mec.es/biosfera/alumno/1bachillerato/petrogeneticos/actividad9/actividad9.htm

Actividad:
http://recursos.cnice.mec.es/biosfera/alumno/1bachillerato/petrogeneticos/actividad9/actividad9.htm
SEDIMENTACIÓN
La sedimentación es la acumulación por deposición de todos aquellos materiales alterados y transportados previamente. Siempre tiene lugar cuando disminuye la energía de los agentes de transporte. Por ejemplo cuando el río llega al mar. Los sitios donde se acumulan los sedimentos se llaman cuencas sedimentarias.
La sedimentación suele producirse con una ordenación de los materiales depositados, desarrollandose lo que se conoce como estructuras sedimentarias. El tipo de estructuras sedimentarias que se generan va a depender de varios factores como pueden ser el agente geológico que produce este proceso, y las características físicas (velocidad de la corriente, dirección de la misma, etc, químicas (presencia de oxígeno, concentración de sales)y biológicas (tipos de organismos presentes, etc) del lugar donde se produce la sedimentación.
El estudio de los sedimentos antiguos (los que encontramos en las rocas sedimentarias) nos permite deducir, por tanto, las condiciones físicas, químicas y biológicas en las que se generaron estos sedimentos cuando las comparamos con las condiciones en las que ocurren en la actualidad.
Algunas de estas estructuras son:
-Estratificación: disposición de los estratos en capas:

Laminación: disposición de finas capas de sedimentos.


Ripple mark: rizaduras producidas sobre un fondo arenoso por corrientes o por olas.


Hay muchísimas más estructuras sedimentarias y otra serie de datos (color, composición) de los sedimentos que nos ayudan a estudiar las condiciones en los que se generaron y reconstruir de esa forma las condiciones ambientales que habia en el pasado. Si queréis saber más esta presentación en power point es muy didáctica:
Suscribirse a:
Comentarios (Atom)
